DIFERENTES TIPOS DE SPLICEOSOME
El
espliceosoma es una maquinaria extremadamente dinámica formada por RNA y proteínas. Existen 5 ribonucleoproteínas
pequeñas (snRNPs) que se ensamblan secuencialmente
en el intrón: primero se unen U1 y U2, depués se ensambla con las anteriores el complejo tri-snRNP formado
por U4, U6 y U5. Una vez formada y ensamblada
al completo, la maquinaria de splicing no esta catalíticamente activa: deben desprenderse las snRNPs U1 y U4 para poder
retirar el intrón en dos pasos consecutivos.
Una
descripción detallada del proceso de splicing resalta la importancia de la
formación de estructuras secundarias en
los duplex de RNA que se forman entre el pre-mRNA y las snRNPs, así como la conveniencia de cambios
conformacionales en estas estructuras de RNA para posibilitar los reordenamientos
estructurales necesarios para el correcto desarrollo del proceso de splicing. La
retirada de intrones requiere dos pasos de trans-esterificación interna en los
sitios de splicing 5´y 3´. Para producir el posicionamiento del RNA sustrato, un complejo ribonucleoprotéico (RNP) consistente
en las 5 pequeñas partículas nucleares snRNPs
presentadas anteriormente y numerosas proteínas debe ensamblarse en cada intrón: es el
espliceosoma. El ensamblamiento del espliceosoma comienza con la unión de la
snRNP U1 al sitio de splicing 5´y
la unión de la snRNP U2. Ambos componentes de RNA (U1 y U2) se aparean base con base con las secuencias del pre-mRNA,
consiguiéndose establecer una estructura
secundaria en la que la adenosina que participará en la primera reacción de trans-esterificación sobresale de la doble
hélice de RNA formada por U2 y el pre-mRNA.
La
adición del complejo tri-snRNP U4-U6-U5
conlleva numerosas reorganizaciones de las interacciones RNA-RNA y RNA-proteínas. El
siguiente paso es el desenrollamiento de U4 de U6 y el apareaminto base con base de U6 con
U2 y con el extremo 5´del intrón,
desplazando a la snRNP U1. De esta forma han quedado fuera del complejo U1 y U4, quedando el espliceosoma catalíticamente
activo: el 2´-OH de la adenosina que sobresale
queda posicionada en el espacio de forma adecuada para realizar el ataque nucleofílico al extremo 5´ del intrón. Tienen que ocurrir ciertas reorganizaciones para acomodar a la pareja de la segunda
trans-esterificación: la snRNP U5 ayuda
en el posicionamiento correcto del extremo 3´-OH del exón 1 para el ataque nucleofílico al exón 2. Tras ligarse los
exones y la liberación del mRNA y del lazo intrónico, el espliceosoma se desensambla y se
vuelve a ensamblar para realizar una nueva
ronda de splicing.
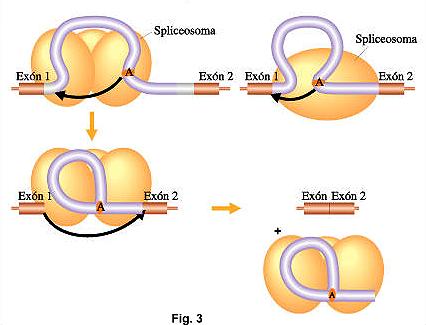
El trabajo
del corte y empalme esta catalizado por una estructura pequeña, compuesta por
ribonucleoproteinas nucleares llamadas snRNPs, constituidas por pequeños
ARN nucleares (snARNs) asociado a proteínas. Su nombre es spliceosoma.
Esta estructura tiene a su cargo el reconocimiento de las secuencias mencionadas
anteriormente en los intrones y su posterior fijación. Luego se desarrollan una
secuencia de pasos que determinan el clivaje y ligado de los intrones y exones.

EXISTEN TRES TIPOS DE SPLICING
Para la eliminación de los intrones antes de la Traducción, los mRNA deben atravesar un proceso denominado Splicing. El término exacto en castellano sería ayuntamiento (gerundio de ayuntar, que es formar una soga nueva al entretejer de los cabos de otras dos), sin embargo el término que ha cuajado es el anglófono, sobre todo porque ya tiene un sentido de especificidad bastante marcado.
El Splicing, esencialmente, es un fenómeno que ocurre entre uno de los extremos (el anterior o 5') del intrón y una posición interna de éste en la denominado sitio de ramificación y conjugado con la unión entre el extremo 3'-OH del exón anterior, que queda libre, y el 5' del exón posterior al intrón. El resultado es que los exones quedan ayuntados y el intrón se libera con una forma de lazada o bucle al formarse un enlace entre el extremo 5' y el sitio de ramificación.

Existen por
lo menos tres tipos generales de splicing: 1) splicing mediado por spliceosoma
o nuclear, una maquinaria enzimática compleja parecida a la
de la iniciación de la Transcripción lleva a cabo el plegado del mRNA y
cataliza las reacciones; 2) auto-splicing de tipo I; 3) auto-splicing
de tipo II. La diferencia entre estos dos últimos es la naturaleza de la
estructura tridimensional formada por el propio RNA del intrón a la hora de
catalizar su propia escisión (la de tipo I se denomina del bolsillo
de Guanina por la intervención de este nucleótido y la estructura que se
forma). Las dos formas de auto-splicing son más propias de las arqueas.
Además de esto, puede también tener lugar Trans-splicing, en el que el
exón de un mRNA se une al exón de otro mRNA diferente, como ocurre en los
mRNAs de los tripanosomas (un grupo de parásitos eucariotas, como el que
causa la malaria).
En el
Splicing hay dos implicaciones muy relevantes: en primer lugar la capacidad de
auto-splicing que muestran algunos intrones refiere a la capacidad
autocatalítica y catalítica del RNA, lo que sostiene la hipótesis de que el
papel de las enzimas fue desempeñado originalmente por RNA en forma de
ribozimas y por tanto este ácido nucleico pudo aportar la base para el
desarrollo posterior de las enzimas/proteínas, más estables y eficientes
energéticamente; en segundo lugar, en el splicing nuclear en eucariotas, en
las juntas de unión de exones resultantes de la eliminación de los intrones
se colocan ciertos complejos protéicos (los Exon Junction Complex o
Complejos de Juntura de Exones) que marcan la posición en que se hallaban
los exones después de su eliminación. La función de estos EJCs parece ser la de
asegurar que no se han producido errores en el splicing y que el mRNA
producido genera la proteína correcta. Esto se debe a que los EJCs son retirados
por el ribosoma durante la Traducción; si no hay una señal de terminación
incorrecta, todos ellos habrán sido retirados pero, si no es así y alguno
permanece, se disparará un mecanismo de destrucción (Decaimiento
Mediado por Secuencias Sin Sentido o Nonsense Mediated Decay) para
eliminar este mRNA y que no siga produciendo proteínas alteradas.
BIBLIOGRAFIA
http://thenewbiologist.blogspot.mx/2008/01/fenmenos-asociados-transcripcin.html
No hay comentarios:
Publicar un comentario